线粒体是细胞内的重要细胞器,它不仅是细胞“能量工厂”,提供生命活动所需90%以上的能量来源;而且被称为“生命能量通货”,是整个细胞生命活动的枢纽。线粒体参与细胞分化、细胞生长、信息传递、细胞周期和凋亡等诸多细胞生理过程,调控能量代谢、自由基与氧化应激、炎症反应、自噬、信号转导、钙稳态、生物合成、组织重建及肌肉发育等组织功能,影响免疫力、体力、记忆力、思维活力、情绪等身体机能。此外,线粒体功能缺陷及能量代谢障碍会损伤高耗能组织和器官,并引起损伤、衰老、代谢紊乱、肌肉退化、癌症、糖尿病、心脏病和神经退行性疾病等病理变化。
然而,与线粒体重要且广泛的生理功能不匹配的是,至今鲜有其在肌肤微观领域,尤其是应用层面及相关活性物研究。线粒体对于肌肤的作用主要体现在其对肌肤老化及肌肤修复两大层面的影响,限于篇幅,笔者将分上下两篇分别阐述。本篇重点讨论线粒体与肌肤衰老的关系。
线粒体与诺贝尔奖
1857年瑞士解剖及生理学家阿尔伯特•冯•科立克在肌肉细胞中发现了颗粒状结构。德国病理及组织学家理查德•阿尔特曼将其命名为bioblast(原生粒)并于1886年发明一种鉴别染色法。1898年德国科学家卡尔•本达用希腊语mitos(线)和chondros(颗粒)组合命名为mitochondrion(线粒体)。1900年美国化学家莱昂诺尔•米歇利斯开发出一种线粒体染色法,并推断其参与氧化反应。英国生物学家大卫•基林在1923~1933年间研究线粒体氧化还原链并辨别出电子载体细胞色素。1931年德国生理学家奥托•瓦尔堡以关于线粒体呼吸酶的研究获诺贝尔生理学医学奖,并于1937~1938年间阐明磷酸三碳糖氧化与形成ATP相偶联的机理。1937年英籍德裔生物化学家汉斯·克雷布斯因提出线粒体三羧酸循环荣获诺贝尔生理学医学奖。1946年比利时生物学家阿尔伯特•克劳德推断负责呼吸链的细胞色素氧化酶位于线粒体。1957年合成出ATP的英国化学家亚历山大•托德,1978年建立化学渗透理论公式及线粒体能量转换机理的英国生物化学家彼得·米切尔,1997年阐明ATP合成酶催化机理和发现离子传输酶的美国保罗•博耶、英国约翰•沃克、丹麦延斯•斯科三位科学家分别获得诺贝尔化学奖。而2019年美国科学家威廉•凯林、格雷格•塞门扎以及英国科学家彼得•拉特克利夫因在细胞感知及对氧气供应适应以及HIF-1(缺氧诱导因子-1)调控线粒体呼吸方面的发现荣获诺贝尔生理学医学奖。
线粒体结构与功能
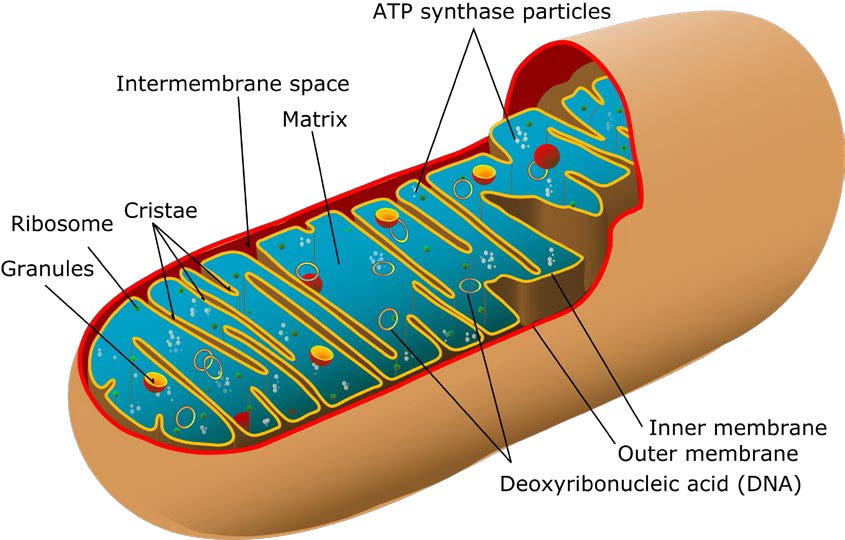
线粒体通常分布在细胞功能旺盛的区域,细胞如肝脏细胞一般含有几百至几千个线粒体,极度耗能组织如心、脑、视网膜中可达1万个,卵子中可高达10万个。线粒体多呈短棒状或长椭圆状,直径约0.5~1.0微米,长1.5~3.0微米,是一个双层膜的囊状小体,由外膜、膜间隙、内膜、基质组成。外膜光滑连续,结构简单,具有通透性,含有孔蛋白如线粒体融合蛋白2(Mfn2),便于胞浆和内膜间物质转运;内膜高度折叠成嵴,内含氧化磷酸化酶复合体和电子传递链;外膜与内膜间的空腔称为外室;由内膜围成的腔称为内室或线粒体基质,内室包含线粒体DNA(mtDNA)和氧化呼吸链的各元件。组成线粒体的结构除了参与合成ATP,为细胞供能外,还承担很多重要的生理功能,如:1)线粒体外膜上的跨膜蛋白Mfn2调节细胞增殖、凋亡和自噬;2)线粒体内膜上的蛋白复合体线粒体膜通透性转换孔(MPTP)影响细胞凋亡和坏死环节;3)线粒体嵴上有许多排列规则的颗粒称为线粒体基粒,存在合成ATP的ATP合酶,其重构可调控线粒体超复合物蛋白等多种内部蛋白的组装。线粒体这些结构的功能证实了线粒体与细胞生命息息相关,并在机体皮肤老化及损伤修复中发挥着重要作用。
人类线粒体DNA(mtDNA)存在于线粒体基质中,是双链的超螺旋环状分子,一般没有内含子,由16569个碱基对构成,是独立于核DNA(nDNA)外的唯一基因组DNA,具有母系遗传、内共生来源、多拷贝性、异质性与纯质性、有阈值效应、高突变性等特点,因此线粒体被称为半自主细胞器。mtDNA包含有37个基因,编码12S和16S两种rRNA、20种tRNA及13个与线粒体氧化磷酸化(OXPHOS)有关的蛋白质,这些蛋白质在有氧呼吸及电子传递链中发挥核心作用。
此外,线粒体可通过融合和裂变来调节自身的形态和功能。融合允许线粒体相互联系,有利于mtDNA信号传递和交换;裂变则确保子细胞器间分离,及时清除功能缺陷的细胞器。
能量代谢与电子传递链
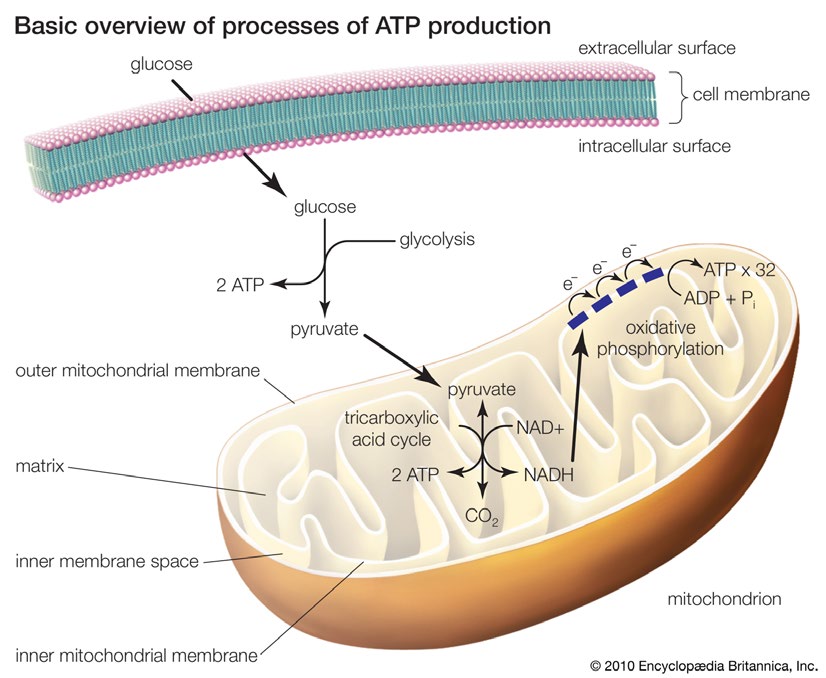
线粒体是真核生物进行能量代谢的结构,以及糖类、脂肪和氨基酸氧化释放能量的场所。以葡萄糖为例,有氧呼吸主要分为三步:1)在细胞质基质中发生糖酵解;2)在线粒体基质中完成三羧酸循环,产生8个还原型烟酰胺腺嘌呤二核苷酸(NADH)和2个还原型黄素腺嘌呤二核苷酸(FADH2)等高能分子;3)通过氧化磷酸化(OXPHOS)利用这些高能分子还原氧气释放能量,合成三磷酸腺苷(ATP)。1分子葡萄糖完全燃烧可释放2870.23kJ/mol的热量,产生30或32个ATP,其中只有2个在线粒体外的细胞质产生,其他都在线粒体产生。
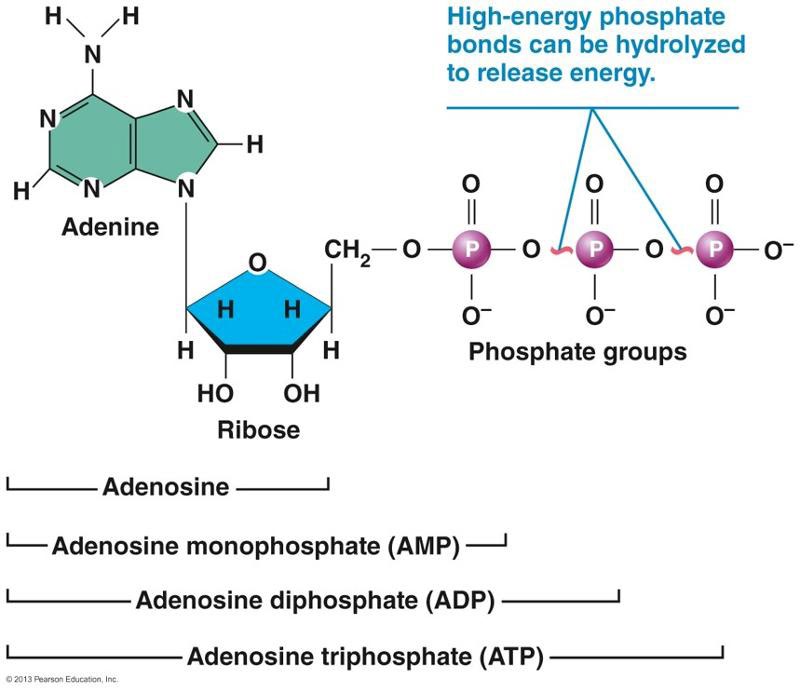
ATP是细胞的直接能量来源,分子简式A—P~P~P,由腺苷、核糖和三个磷酸基团组成,分子量507.184。ATP含有两个高能磷酸键,极不稳定,可水解释放30.54kJ/mol能量,与二磷酸腺苷(ADP)相互迅速转化实现贮能和放能平衡,从而保证生命活动的能量供应;脱两个磷酸生成单磷酸腺苷(AMP),为RNA的基本组成单位。据估算,线粒体每秒可生产600个ATP,细胞每分钟可产生30亿个ATP。人体中ATP瞬时含量仅约50.7克,只能维持0.3秒剧烈运动,但每天可循环制造50~75kg的ATP,相当于成人的自身体重。
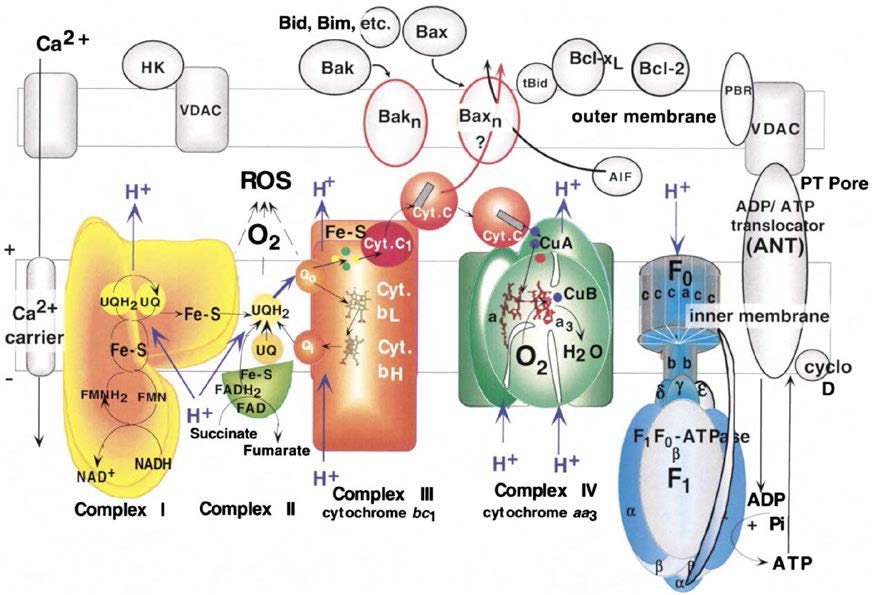
电子传递链(ETC)指线粒体在有氧呼吸过程中的电子转移反应,因其与呼吸作用直接相关,也被称作呼吸链。电子传递链主要由位于线粒体内膜上的4种酶复合物I(NADH脱氢酶)、II(琥珀酸脱氢酶)、III(细胞色素C还原酶)、IV(细胞色素C氧化酶)和2种可移动电子载体辅酶Q(CoQ,又称泛醌UQ,介于Ⅰ/Ⅱ与Ⅲ之间)及细胞色素C(CytC,介于Ⅲ与Ⅳ之间)构成。以这些复合物为基础,可以聚合形成多层次的呼吸链超级复合物乃至超超级复合物,从而获得更高的稳定性和更高的能量转换效率。营养物质氧化产生的NADH和FADH2将电子与质子转移到线粒体膜间隙中,积累电化学势差,产生质子流,进而制造ATP。
活性氧自由基与氧化应激
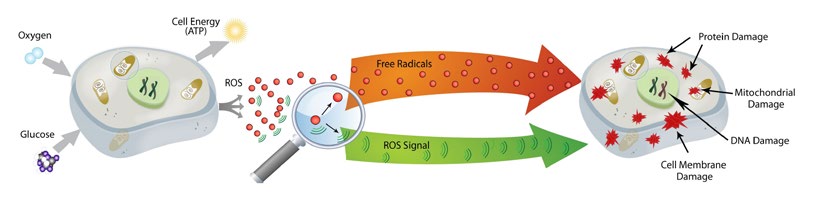
线粒体直接利用氧气(O2)制造能量,大多数细胞中超过90%的O2在线粒体中消耗。而电子伴随NADH和FADH2在高氧环境和高还原态的呼吸链中总会泄露,尤其在复合体IV中,大量电子从线粒体内膜和基质中漏出,将大约2%的O2还原为水(H2O),产生多种氧的单电子还原产物,如一电子还原产物超氧阴离子(O2•-)、二电子还原产物过氧化氢(H2O2)、三电子还原产物羟基自由基(•OH)等,统称活性氧(ROS)。同时氮气(N2)也参与产生少量活性氮(RNS)。电子的损失被称为氧化,线粒体呼吸链中的电子传递是ROS的主要发生器,作为氧化产物的ROS和RNS就是自由基。
过量ROS与自由基泄露到线粒体外进入细胞及胞外,将电子传递给其他分子,对人体造成的氧化损害,称为氧化应激(OS)。广义的氧化应激指机体在遭受有害刺激时,ROS的产生和清除失去平衡,引起炎性细胞侵入所造成的组织损伤。氧化应激具有重要的生理功能,如调节体内氧供需平衡、突触可塑性、抗菌防御反应、炎症、学习和记忆等,可通过能量衰竭、DNA氧化损伤、脂质过氧化、线粒体自噬与功能障碍、细胞损伤与凋亡等途径起作用。
事实上,线粒体正常代谢产生的适量ROS是健康且对机体有利的。例如吞噬细胞在细胞膜受到刺激时,通过呼吸暴发机制产生的大量ROS是其发挥吞噬和杀伤作用的主要介质。研究表明ROS可诱导和维持多潜能干细胞多能性、细胞分化和重编,并可通过调控上皮间质转化过程影响肌肤衰老、炎症发生发展进程与表皮修复。
过量摄入外源性抗氧化剂(如VC、VE、视黄醇、番茄红素、虾青素)不仅对身体及肌肤无益,反而有害,因其过多会造成细胞对ROS和自由基信号不敏感,导致低效线粒体无法及时清除,反而造成ATP生产长期不足,缩短细胞寿命,加速肌肤衰老。最重要的是从源头上减少自由基产生,提升线粒体自身抗氧化能力,而非一味补充外源性抗氧化剂。
线粒体功能损伤与衰老
细胞衰老指细胞在生命活动中增殖分化能力和生理功能完整性逐渐退化的过程。衰老的特点有呼吸能力下降、能量代谢减少、氧化应激增加、蛋白质氧化、DNA损伤突变、脂质过氧化、自噬障碍、氧化磷酸化障碍、干细胞稳态变化等。研究发现众多疾病与细胞衰老有关,如肿瘤、特发性肺纤维化、心血管疾病、神经退行性疾病、糖尿病等。肌肤老化是机体衰老的一部分,而衰老进程在皮肤上表现得最明显。近年来对衰老分子机制的研究主要有氧化自由基、DNA损伤积累和端粒缩短等学说。
传统自由基损伤理论认为高水平ROS具有细胞毒性,作为能量代谢副产物影响线粒体功能,造成细胞脂质、蛋白质、DNA损伤,导致细胞衰老。1950年Harmon首次提出ROS直接调控细胞衰老过程,同时Linnane和Fleming认为过量ROS导致mtDNA突变,引发细胞衰老。线粒体质量、活性改变及电子传递链复合体的生物合成直接影响细胞衰老,线粒体损伤及功能障碍是衰老的始发环节,且主要与ROS代谢有关。研究也表明间充质干细胞及成纤维细胞线粒体ROS的清除及其稳态的维持能有效抑制细胞衰老。
此外,线粒体形态、结构、数量的变化也与细胞衰老密切相关,主要表现为线粒体体积肿大、数目增减、内膜损伤、嵴缩短及线粒体变性导致空泡和破裂等。这些改变受到线粒体生成和自噬相关基因的协同调节,稳态失衡导致功能障碍线粒体的累积,造成细胞功能衰退。同时,线粒体动力学的改变对细胞衰老也发挥重要作用,线粒体的形态和功能可通过融合和裂变来调节。融合允许线粒体相互联系,有利于mtDNA信号传递和交换;裂变则确保子细胞器间分离,及时清除功能受损的线粒体,减少ROS的过量产生和氧化损伤。
mtDNA突变与衰老
mtDNA突变及累积在人体衰老中的作用已被广泛证实,且被认为是肌肤老化尤其是皮肤光老化的重要因素。皮肤组织mtDNA突变的特点主要有:1)mtDNA缺乏组蛋白和内含子,极易发生突变;2)存在多重皮肤与衰老相关突变,如点突变、缺失、重排、插入和小串联重复;3)突变随年龄增高累积;3)突变在皮肤组织中不均衡分布,以真皮成纤维细胞及毛囊组织居多;4)皮肤组织中突变累积与环境因素相关,以ROS自由基最重要,主要由长期反复高强度紫外线、离子辐射引起的DNA链断裂导致,且皮肤暴露部位突变发生率和丰度高于非暴露部位;5)其他外界因素包括吸烟、环境及食物中的化学因素导致的核苷酸烷基化,X线与化学性致癌剂导致的DNA交联等;6)内部因素有DNA聚合酶错误掺入、mtDNA内在热不稳定性引起的自发化学突变、缺乏组蛋白保护及相应的修复系统等。
mtDNA突变后所产生的生物学效应有两种模式:细胞生物能阈值模式和细胞程序性死亡模式。发生在mtDNA结构基因上的突变大多与细胞呼吸功能有关,一方面造成线粒体呼吸相关功能组分的合成障碍和缺失,导致线粒体呼吸能力下降,引发细胞衰老;另一方面造成mtDNA复制与修复障碍,通过选择性扩增和累积,产生呼吸链局部障碍,加重线粒体损伤,导致生物能缺陷,加剧细胞衰老。其中复合体Ⅰ受影响最大。最新研究表明,mtDNA敲除鼠几周内皮肤明显起皱、大面积掉毛发;逆转mtDNA突变恢复线粒体功能后,小鼠皮肤皱纹消失、毛发重新生长,已有衰老症状的小鼠“返老还童”,表明线粒体是皮肤衰老起皱、脱发的可逆调节因子。
炎症反应与衰老
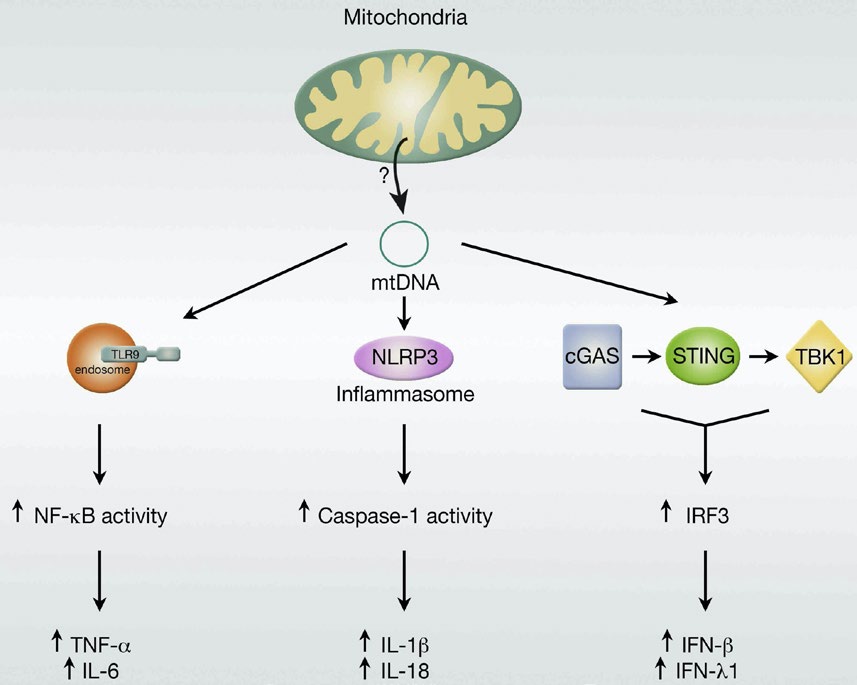
线粒体的代谢、稳态改变、自嗜、凋亡过程均与炎性体的激活密切有关,影响细胞及肌肤老化进程。衰老细胞会产生一系列分泌因子,包括基质金属蛋白酶(MMP)、生长因子(GF)和促炎症细胞因子,统称衰老相关分泌表型(SASP)。SASP的形成与线粒体损伤调节的免疫应答的激活过程密切相关,可加重衰老细胞的促衰老效应。mtDNA作为细胞损伤相关分子模式(DAMPs)之一,是线粒体损伤与炎症功能间联系的标志物。一方面,mtDNA可通过结合并激活三条炎症通路的相应受体,即Toll样受体蛋白(TLR)、核苷酸结合寡聚化结构域样受体蛋白(NLRP)和细胞质循环GMP-AMP合成酶-干扰素基因刺激因子DNA(cGAS-STINGDNA)传感系统介导的途径参与炎症反应,同时损伤的线粒体可以释放甲酰多肽,通过甲酰多肽受体1(FPR1)激起免疫应答;另一方面,线粒体功能与ROS代谢异常同样可以激发炎症反应。尽管如此,在细胞衰老过程中,线粒体损伤既可能是炎症的激发剂,也可能是其结果,均可一定程度促进衰老发生与发展。持续炎症刺激可激活循环免疫细胞,在循环中释放的细胞因子、趋化因子、NO和ROS可进一步诱导线粒体损伤,反过来通过激活mtDNA诱导炎症途径,形成恶性循环,加强整个过程。
线粒体自噬与衰老
线粒体自噬的概念最早由Lemasters提出,指一种细胞防御代谢及自我修复的过程,是细胞维持体内平衡的极端尝试。在细胞衰老过程中,线粒体稳态失衡会导致损伤线粒体的过度增殖以及细胞功能衰退,通过选择性清除衰老、损伤及功能失调的线粒体,可促进其更新并维持其功能。在整体细胞器退化损伤之前,线粒体可以采取囊泡内消除这一替代途径处置受损细胞器;若线粒体受损严重则通过线粒体裂变等动力学机制触发其断裂和降解,最终被消除。目前融合-裂变紊乱被认为是在应激或衰老过程中线粒体结构异常和功能失调的机制之一,研究发现线粒体融合和裂变异常可引发异常线粒体聚集,导致线粒体自噬受损。提高衰老细胞中线粒体自噬水平有助于清除损伤线粒体,降低细胞内ROS,缓解细胞衰老表型,从而保持细胞活性。
线粒体与干细胞衰老
干细胞对人体自我修复和组织再生至关重要,干细胞衰老通常伴随组织代谢和再生潜能下降等衰老迹象,且与线粒体功能衰退密切相关。研究表明ROS作为信号分子,对干细胞增殖分化有调节作用,高水平ROS对干细胞功能具有损伤作用。对mtDNA突变鼠给予抗氧化剂治疗,其存在的干细胞缺陷及功能损伤得到改善,表明由线粒体异常引发的ROS代谢异常可直接干扰并损伤小鼠干细胞功能。线粒体维持干细胞功能的另一种机制是通过调节代谢物乙酰辅酶A(AcetylCoA)、烟酰胺腺嘌呤二核苷酸(NAD+)、腺嘌呤核糖核苷酸(AMP)、S-腺苷甲硫氨酸/S-腺苷同型半胱氨酸(SAM/SAH)的比率来调节干细胞的自我更新和稳态,同时干细胞可以通过一定机制清除功能缺陷的线粒体。实验证实通过线粒体自噬对线粒体质量的控制可以有效清除干细胞中功能缺陷的线粒体,维持干细胞功能。
结语
线粒体的功能完整性与肌肤健康密切相关,本文作为上篇对线粒体发现与研究的历史、结构功能、能量代谢与电子传递链、活性氧自由基与氧化应激等概念做了简要梳理,并重点讨论了ROS代谢、mtDNA突变、炎症反应、线粒体自噬及干细胞功能调节与细胞及肌肤衰老的关系。笔者将在下篇进一步讨论线粒体对组织再生、损伤修复、炎症反应的激活,进而影响肌肤修复的策略。遗憾的是,在护肤领域,目前鲜有以线粒体为靶点进行肌肤抗衰及肌肤修复的相关研究,能够同时兼顾肌肤修复与抗衰需求的全能功效成分更是少之又少。但随着中国消费升级大潮的来临及精准护肤概念的推进,加之市场与成分党对靶标明确、安全有效功能护肤成分的热衷与呼唤,相信线粒体作为护肤领域新的金矿,将引起更多业内科研人员、工程师、品牌商乃至消费者的青睐。
参考文献:
[1]NewmeyerDD.etal.Mitochondria:releasingpowerforlifeandunleashingthemachineriesofdeath.Cell2003:112:481.
[2]BalabanRS.etal.Mitochondria,oxidants,andaging.Cell2005,120:483.
[3]OttM.etal.Mitochondria,oxidativestressandcelldeath.Apoptosis2007,12:913.
[4]MurphyMP.Howmitochondriaproducereactiveoxygenspecies.Biochem.J.2009,417(1):1.
[5]PanelM.etal.Mitochondriaandaging:aroleforthemitochondrialtransitionpore?AgingCell2018,Articlee12793.
[6]SinghB.etal.Reversingwrinkledskinandhairlossinmicebyrestoringmitochondrialfunction.CellDeathDis.2018,9:735.
[7]StoutR.etal.Mitochondria’sroleinskinageing.Biology2019,8:29.
彗搏科技CEO卢云宇博士
 供需大厅
供需大厅
 登录/注册
登录/注册 供应商登录
供应商登录