绪言
解决脱发所带来的烦恼正在成为一大刚需。关于脱发问题的严重性目前有各种报道,包括:“中国平均6个人中就有1个有脱发症状……”,“中国脱发人群已超2亿(2.5亿)……”,“77%有脱发困扰……”,“90后成为防脱消费的主力军……”,等等。解决脱发问题的可能方法包括:1)药物治疗:以米诺地尔为首的外用药对10%左右的患者有显著效果。2)植发:将自身的毛发通过移植来再分配,但毛发总数量没有增加。3)毛发再生医疗:目前利用干细胞的毛发再生医疗已进入临床II期研究阶段,期待不久的将来可以实现临床治疗。4)防脱发:属于预防为主的大健康思路,理论上讲可以通过特殊化妆品来而实现。其中防脱发是化妆品企业应该重点关注的领域。
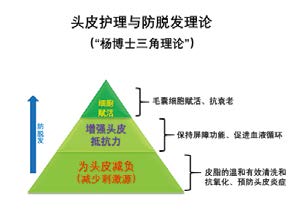
保持头皮的健康是预防由于头皮炎症而引发的脱发之基础,防脱发则是头皮护理的终极目标。图1描述了头皮护理的基本技术开发方向,包括三个层面,即为头皮减负﹑增强头皮抵抗力﹑毛囊细胞赋活。作为其中最高层面的毛囊细胞赋活,传统方法是通过有效成分,促进有利于头发生长的生长因子如FGF﹑BMP等;或者抑制不利于头发生长的生长因子,如TGF-b﹑NT-4等。本文通过在对近期不同学者的学术论文分析与归纳的基础上,提出毛囊细胞赋活的新模式,即通过提升毛囊干细胞的自噬活性,而实现头皮抗衰老与防脱发。
细胞自噬与自噬活性
自噬(Autophagy)一词源自希腊词语auto(自我)及Phagein(吃),意为“将自己吃掉”。1963年,比利时科学家Christiande Duve因发现细胞能够将自身成分用膜包起来,形成袋状囊泡并运送到溶酶体(Lysosome)降解掉而发明了Autophagy一词,也因发现这种新型的细胞器─溶酶体而获得了1974年诺贝尔生理学或医学奖[1]。自噬作为细胞内的再循环系统(recyclingsystem),其作用主要是清除降解细胞内受损伤的细胞结构、衰老的细胞器、以及不再需要的生物大分子等。自噬作用在消化的同时,也为细胞内细胞器的构建提供原料,即细胞结构的再循环。自噬作用机制失灵将导致细胞异常甚至死亡,其功能下降与癌症、神经变性疾患、感染性疾患、心血管疾患、代谢疾患、肺疾患、生活习惯病等各种疾病,以及老化、寿命延长等有关[2]。细胞自噬机制的关键因素是自噬基因,自噬基因的稳定性直接影响到细胞自噬功能[3]。2016年东京工业大学名誉教授大隅良典因在细胞自噬机制方面的发现,获得当年的获诺贝尔生理学或医学奖。[4]如今自噬作用及其医学临床应用的相关研究正在迎来爆发性的发展期。近年来,自噬调控的应用也成为化妆品研究领域的一个新热点。如皮肤的抗衰老、美白及防脱育发等。
根据细胞内底物进入自溶酶体腔的方式不同,细胞自噬可分为巨自噬(macroautophagy)、微自噬(microautophagy)和分子伴侣介导的自噬(chaperonemediated autophagy,CMA)三种方式[2]。单纯说自噬一般指的是巨自噬,它包含4个连续过程:1)诱导内质网内生成的双层膜构成的隔离膜包裹吸收部分细胞质出现吞噬泡(phagophore),其特征为:新月状或杯状,双层或多层膜,有包绕胞浆成分的趋势;2)自噬体组装,组装完成后形成自噬体(autophagosome)的特征为:双层或多层膜的液泡状结构,内含胞浆成分,如线粒体、内质网、核糖体等;3)自噬体与带有加水分解酶的溶酶体融合形成自噬溶酶体(autophagolysosome);4)降解和降解物质的循环再利用。自溶酶体内的各种大分子在水解酶的作用下,可被分解为小分子物质。例如,能将蛋白质分解为多肽或游离氨基酸;把核酸分解为核苷和磷酸;将碳水化合物分解为寡糖类或单糖;将中性脂肪分解为甘油和脂肪酸等。这些被分解而生成的可溶性小分子物质,能透过溶酶体体膜,重新参与细胞的物质代谢,一些未被完全消化的物质残留下来,形成残余小体。经消化内含物质生成的自体溶酶体或称为自噬泡(autolysosome)其特征为:单层膜,胞浆成分已降解[5]。
细胞正常情况下很少发生自噬,除非有诱发因素的存在。诱发因素很多,既有来自于细胞外的,如外界中的营养成分、缺血缺氧、生长因子的浓度等;也有细胞内的诱发因素,如代谢压力、衰老或破损的细胞器、折叠错误的蛋白质,DNA损伤等。这类诱发因素的转递路径被称为自噬信号通路。法国科学家Patrice Codogno博士于2000年首先证实了PI3K信号通路在自噬的作用,I型抗自噬,III型促自噬,是自噬信号通路的开拓者[6]。目前的研究发现,自噬调节涉及多种信号通路,其中以腺苷单磷酸活化蛋白激酶AMPK(adenosine mono-phosphate activated protein kinase)及哺乳动物雷帕霉素受体mTOR(mammaliantargetofrapamycin)信号通路为调控核心。AMPK促进自噬发生,而mTOR抑制自噬发生。如图2所示。
自噬起始最为关键的环节之一是自噬小体(autophagosome)的形成。自噬标志性蛋白LC3不仅参与自噬小体的形成,也是自噬途径的底物受体,在自噬过程中起着至关重要的作用。自噬小体形成时,胞浆型LC3-I被半胱氨酸蛋白酶切去掉一小段多肽的LC3被称作胞浆型LC3-I,与自噬体的膜构成成分的磷脂酰乙醇胺结合后成为膜型LC3-II。这样,LC3-II/I比值的大小可估计自噬水平的高低。自噬体一旦与溶酶体融合LC3-Ⅱ就会被分解。如果使用溶酶体阻碍剂处理,LC3-Ⅱ就不会被分解地蓄积下来,所以通过测定添加了溶酶体阻碍剂的样品与未添加的样品的LC3-Ⅱ量得到的差值可以被认为是因自噬被分解的LC3-Ⅱ量、即自噬通量。自噬通量是自噬活性的指标,该量值越大自噬活性就越高。p62(SQSTM1/sequestosome1)是和LC3有关的自噬相关蛋白,通过与LC3结合被自噬体选择性吸收分解,因此,细胞内的p62蛋白量变与自噬活性呈负相关。
调查自噬活性的方法有定量自噬体的数量的方法和定量自噬通量(Autophagyflux)的方法。自噬体的形成可根据其形态特征(双层膜)用透射电子显微镜调查,或者将自噬相关标记蛋白如LC3免疫染色后用荧光显微镜观察、计算辉点。由于不能辨别是因诱导了自噬而使自噬体增加了还是自噬的最终分解阶段被阻碍而使自噬体增加了,自噬体数量的变化不能代表自噬的活性,自噬活性大多是通过定量相关蛋白自噬通量来确定的。
毛嚢结构﹑毛发生长周期与防脱育发
头皮是头发生长的土壤。毛发源自表皮中的深囊状结构—毛囊。毛囊是指包围头发(毛干)的组织层,以发丝为中心呈同心圆状的层构造。靠近发丝的内侧由上皮性成分(内毛根鞘、外毛根鞘)构成,外侧由结缔组织性成分(结缔组织毛囊)构成。毛根向下延伸到毛囊中,并在其底部变宽形成内缩的球被叫做毛球、中央有毛乳头。毛母细胞是为了包围毛乳头而存在的细胞,毛母细胞分化分别形成毛干及内毛根鞘、外毛根鞘。毛母细胞中也混杂着生成黑色素的色素细胞(黑色素细胞)。毛乳头细胞中释放出众多因子、会给毛母细胞带来影响(增殖及分化)。
通常情况下,毛发的生长呈现周期性,分为生长期(anagen)、退行期(catagen)、休止期(telogen)[7,8]。头发持续成长2-6年(成长期)、之后历经2-3周衰退(衰退期)、停滞3-4个月(休止期)。当休止阶段结束时,老的毛发就会脱落。然后毛囊回到生长期,且新的毛发开始生长。健康头皮的头发中约85%处于成长期、约1-2%为衰退期、约15%为休止期。每人平均十万根头发,以生长周期5年计算,则每年的自然脱发数为100000÷5=20000,即两万根。而折合到每天的自然脱发数则应为20000÷365≈55根。而如果按生长周期为4年来计算的话,则每年自然脱发数为25000根,每天约为68根。因此,每天脱落50~100根都是正常的。而远超这个数量则属于异常。从统计学的观点看,脱发是处于不同生长期毛囊量比变化的结果,一旦处于休止期的毛囊占比增加就会导致脱发,因此延长生长期缩短休止期可以防止脱发。
自噬活性与毛发生长
毛发生长是一个复杂的生理过程。从毛囊干细胞活化增殖、到有核细胞向无核、无细胞质的结构细胞的转变是毛发产生的关键。这种形式的细胞“死亡”或角质化,需要去除细胞器,使细胞质充满角质蛋白束,而角质蛋白束则需要进一步交联才能形成浓密的毛发。JonesLA等描述了这种形式的细胞“死亡”过程,其中包含细胞凋亡,自噬,线粒体自噬等方式[9]。
相对于程序性“死亡”的细胞凋亡,自噬作为降解和回收细胞成分的基本过程,在毛发生长中的作用仍然存在争议。一方面,由于来自自噬相关基因7(ATG7)缺陷小鼠的皮肤移植表现出毛发生长异常,毛发生长可能需要自噬[10]。另一方面,心理压力导致自噬和头发周期延迟[11]。
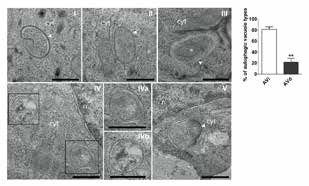
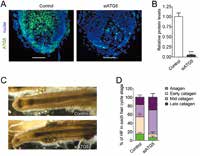
Parodi.C[12]等以人类头皮毛囊作为一个(微型)器官研究模型,经器官培养后制作成的冻结切片,在透射电子显微镜(TEM:transmission electron microscopy)下观察到不同生长阶段毛母细胞自噬体(图2)。进一步定量不同生长期的头皮毛囊细胞自噬体的数量及数量变化,及不同条件下相关自噬标志蛋白的表达(LC3B和p62的自噬通量)调查了自噬活性和毛发生长周期的关系。
在透射电子显微镜下观察时,可以看到在成长期毛囊中毛母细胞中有很多带有是自噬体的特征的双层膜构造的囊泡、以及隔离膜渐渐吸收分解物形成自噬体的过程(图3Ⅰ~Ⅲ。可以看出双层膜形成的状况)。在初期的自噬体(AVi:initial autophagic vacuoles)中可以观察到形态上尚未被分解的核糖体及线粒体等细胞内器官(图3Ⅲ)。在后期的自噬体(AVd:degradative autophagi cvacuoles、自噬溶酶体)中内含物质已经被分解(图3Ⅳ、Ⅴ)。调查成长期毛囊中AVi和AVd的比例,发现AVi较多(图3)、看不到AVd的蓄积,由此可以看出成长期毛母细胞中自噬较为活跃。
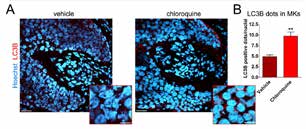
通过共聚焦显微镜观察发现在毛乳头附近的毛母细胞(proximal hair matrix、增殖最猛烈的部分)及分化成内毛根鞘及毛干前的毛母细胞(precorticalhairmatrix)中有大量LC3B(LC3-Ⅱ)。一旦对成长期毛囊使用是自噬阻碍剂(阻碍溶酶体和自噬体融合)毛母细胞中表达的LC3B就会增加(图3A,B)。进一步表明成长期毛母细胞中自噬较为活跃。
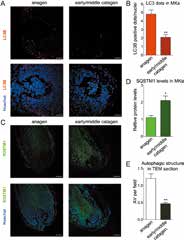
通过比较处于成长期(第Ⅳ阶段)及衰退期(初期~中期)的人体头皮毛囊中的毛母细胞中的自噬发现:在衰退期LC3-Ⅱ的量相对减少了(图4A,B)、另一方面,在衰退期相应的p62残留较多(图4C,D),该结果说明毛母细胞在衰退期的自噬活性比在成长期的较低。另外在TEM观察中,与成长期相比,衰退期的自噬泡(AutophagicVacuoles)数量较少(图4E),进一步确认了在衰退期自噬活性相对较低。
Parodi.C等进一步调查了抑制自噬对毛发周期的影响。一旦导入siRNA(small interference RNA),自噬关联基因ATG5的基因表达被抑制(图5A,B)、LC3-Ⅱ减少、p62增加了,表明毛囊的自噬因转染siATG5而下降。然后从形态学、免疫组织学方面调查96小时后的毛发所处生长周期。没有转染的对照组毛囊是从成长期缓慢地转移到衰退期的,而转染了siATG5(即抑制了自噬)的毛囊,从成长期到衰退期的转移相对更快,许多毛囊显示处于从衰退期中期到末期的阶段、而停留在成长期(第Ⅳ阶段)的毛囊仅10%以下(图5C,D)。
既然抑制毛囊细胞自噬就会促使毛囊生长更早进入衰退期,那么激活毛囊细胞的自噬是否可以延长毛囊生长期呢?
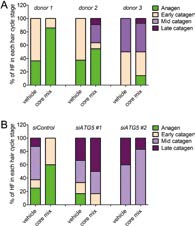
Parodi.C等人用含有人们熟知的自噬促进剂亚精胺类化合物[13~16]的市售防脱发产品(coremix)诱导毛囊细胞自噬。对成长期毛囊(第Ⅳ阶段)添加自噬诱导剂coremix培养5天调查毛发周期,与对照(vehicle)相比处于成长期的比例更高(图6A)。虽然在培养期的末尾(第5天)成长期(第Ⅳ阶段)毛囊正在进入衰退期,但通过自噬激活处理约16%(3组试样平均值)保持在了成长期(图6A)。未经siATG5处理的成长期毛囊通过coremix的自噬诱导作用成长期的比例变高了,但经siATG5转染了的毛囊上没能确认到该效果(图6B)。
进一步对存在于Auber临界线以下的毛母细胞的增殖及凋亡分类调查发现:对于Auber临界线以下的毛母细胞,通过转染siATG5,抑制自噬,减少了Ki-67positive细胞(仍在增殖的细胞)的比例、TUNELpositive细胞(发生了凋亡的细胞)增加。也就是说一旦抑制自噬,毛囊的成长期就会提前结束、凋亡增加进入衰退期。对于Auber临界线以下的毛母细胞,通过coremix激活自噬,提高了Ki-67positive细胞(在增殖的细胞)的比例、降低了发生凋亡的细胞的比例。也就是说,一旦激活自噬,毛囊的成长期就会延长。
上述离体人毛囊(ex vivo human hair follicles)培养实验结果表明:成长期毛母细胞的自噬较为活跃,衰退期自噬活性下降,激活毛囊细胞的自噬可以延长毛囊生长周期,自噬对于维持人类毛发的健康生长至关重要。问题是自噬调控是否可以激活休止期毛母细胞。
黄箐等人[17~19]通过活体老鼠自噬调控(invivo)实验,观察了休止期的老鼠经自噬诱导后毛发生长的情况,发现影响ATP三磷酸腺苷合成酶及mTOR信号回路的代谢产物α-KB等可以作为自噬诱导剂,通过激活自噬的方式激活休眠毛囊,并启动新的毛发生长周期。
虽然自噬诱导剂很有可能是清除毛发成长障碍的有效方法,但支配毛发生长周期的因子还有很多。已知的有:角质形成细胞生长因子FGF7(别名KGF:keratinocyte growth factor)在成长期表达显著,可防御UV照射、化学疗法及毒物等导致的细胞死亡;而FGF5会诱导衰退期;生物钟基因(Period1,BMAL1)控制毛发周期,如果敲除Period1,BMAL1成长期就会显著变长[20],且BMAL1与蛋白酶体、自噬两者的活化有关[21,22]。人们对这些因素和自噬的关联性的认识还非常有限。虽然已有把提升自噬活性的咖啡因[23]作为药用化妆品(cosmeceutical)销售,并称可延长成长期、促进毛母细胞的增殖[24]。但现在能够作为化妆品有效成分用于防脱育发的自噬诱导剂还非常有限,相关的研究还处于初级阶段。
参考文献
1. Klionsky DJ, Cueva R, Yaver DS, J Cell Biol. 1992 Oct;119(2):287-99.
2. Mizushima N, Levine B, Cuervo AM, Klionsky DJ, Nature. 2008 Feb 28;451(7182):1069-75.
3. Levine B, Kroemer G, Cell. 2019 Jan 10;176(1-2):11-42.
4. www.nobelprize.org/prizes/medicine/2016/summary/
5. Boya P, Codogno P, Rodriguez-Muela N, Development. 2018 Feb 26;145(4).
6. Petiot A, Ogier-Denis E, Blommaart EF, Meijer AJ, Codogno P, J Biol Chem. 2000 Jan 14;275(2):992-8.
7. Müller-Röver S, Handjiski B, van der Veen C, Eichmüller S, Foitzik K, McKay IA, Stenn KS, Paus R, J Invest Dermatol. 2001 Jul;117(1):3-15.
8. Schneider MR, Schmidt-Ullrich R, Paus R, Curr Biol. 2009 Feb 10; 19(3): 132–142
9. Jones LA, Harland DP, Jarrold BB, Connolly JE, Davis MG, Br J Dermatol. 2018 Jun;178(6):1341-1352.
10. Rossiter H, Stübiger G, Gröger M, König U, Gruber F, Sukseree S, Mlitz V, Buchberger M, Oskolkova O, Bochkov V, Eckhart L, Tschachler E, Exp Dermatol. 2018 Oct;27(10):1142-1151.
11. Wang L, Guo LL, Wang LH, Zhang GX, Shang J, Murao K, Chen DF, Fan XH, Fu WQ, Arch Dermatol Res. 2015 Mar;307(2):171-81.
12. Parodi C, Hardman JA, Allavena G, Marotta R, Catelani T, Bertolini M, Paus R, Grimaldi B, PLoS Biol. 2018 Mar 28;16(3): e2002864.
13. Pietrocola F, Lachkar S, Enot DP, Niso-Santano M, BravoSan Pedro JM, Sica V, Izzo V, Maiuri MC, Madeo F, Mariño G, Kroemer G, Cell Death Differ. 2015 Mar;22(3):509-16.
14. Michiels CF, Kurdi A, Timmermans JP, De Meyer GRY, Martinet W, Atherosclerosis. 2016 Aug; 251:319-327.
15. Bhukel A, Madeo F, Sigrist S, Autophagy. 2017 Feb;13(2):444-445.
16. Chrisam M, Pirozzi M, Castagnaro S, Blaauw B, Polishchuck R, Cecconi F, Grumati P, Bonaldo P, Autophagy. 2015;11(12):2142-52.
17. Fu X, Chin RM, Huang J. et al., Cell Metab. 2015 Sep 1;22(3):508-15.
18. Chai M, Huang J.et al., Cell Rep. 2019 Jun 18;27(12):3413-3421.
19. J. Huang, et al., WO2016003854A1
20. Al-Nuaimi Y, Hardman JA, Bíró T, Paus R. et al, J Invest Dermatol. 2014 Mar;134(3):610-619.
21. Huang G, Zhang F, Ye Q, Wang H, Autophagy. 2016 Aug 2;12(8):1292-309.
22. Jeong K, He B, Nohara K, Park N, Shin Y, Kim S, Shimomura K, Koike N, Yoo SH, Chen Z, Sci Rep. 2015 Jul 31; 5:12801.
23. Mathew TS, Ferris RK, Downs RM, Kinsey ST, Baumgarner BL, Biochem Biophys Res Commun. 2014 Oct 24;453(3):411-8.
24. Fischer TW, Herczeg-Lisztes E, Funk W, Zillikens D, Bíró T, Paus R, Br J Dermatol. 2014 Nov;171(5):1031-43.
高桥元次,甘智斌,杨建中
一般社团法人日中化妆品国际交流协会
美研创新株式会社

*3月荣格网络研讨会推广
 供需大厅
供需大厅
 登录/注册
登录/注册 供应商登录
供应商登录