 供需大厅
供需大厅
 登录/注册
登录/注册 供应商登录
供应商登录










荣格工业资源APP
了解工业圈,从荣格工业资源APP开始。
欢迎来到荣格工业资源网!

供需大厅

登录/注册
公众号

更多资讯,关注微信公众号
小秘书

更多资讯,关注荣格小秘书
邮箱
您可以联系我们 info@ringiertrade.com

电话
您可以拨打热线
+86-21 6289-5533 x 269
建议或意见
+86-20 2885 5256
顶部

荣格工业资源APP
了解工业圈,从荣格工业资源APP开始。



近年来,“头皮微生态”概念已掀起国内外洗护行业的新潮流。据报道,2024–2025年,中国头皮微生态洗护产品市场规模已突破10亿元,年增速高达约 91%,远高于整体洗护品类的约 2.2% 。

然而截至目前,“微生态”概念在洗护市场的渗透率仅 6.7%,远低于护肤的15%以上水平,存在广阔增长空间。核心销售品类为洗发水、套装类、免洗喷雾式洗发水与头皮精华/油类产品,合计占比最高。小红书等社交平台上,“头皮微生态”相关标签阅读量已达数亿,消费者认知迅速提升。大部分精准洗护消费者(80% 以上)已知晓微生态概念,其中超过 70% 的人认可菌群平衡的重要性,57% 认为微生态产品能改善头油、头屑、脱发等问题的根源。
从成分与技术角度,目前市场上的产品逐渐采用益生元(prebiotics)、益生菌(probiotics)和后生元(postbiotics)等组合,调节头皮菌群多样性,缓解炎症与刺激 。同时,洗护产品越来越像“头皮皮肤科”,强调屏障修护、菌群平衡,推出不同头皮类型(如油性、干性、有屑等)的细分方案。国际趋势亦聚焦“头皮皮肤化”(scalpcare → skinification),2025年全球头皮微生态护理将全面加速,成核心关注方向 。
“抑恶扬善”式菌种平衡策略是当前皮肤、肠道等微生态护理领域常提到的一种精准调控菌群的方法,其核心理念是:不是“一刀切”式杀菌,而是针对性抑制有害菌(pathobiont),同时扶持或促进有益菌(commensals/beneficial bacteria)的生长与活性,从而恢复并维持菌群平衡(microbiome balance)。作者于数年前开始提倡并践行抑恶扬善式的头皮菌种平衡的技术开发策略,本文将就其核心概念及理论基础进行阐述。
头皮微生态平衡的定义
皮肤是人体最大的器官,每一单位面积上都栖息着丰富的微生物群落。头皮及其覆盖的毛发形成了一个独特且复杂的微生态系统。头皮微生态(scalp microecology)是指定植于头皮表面的多种微生物(主要包括细菌、真菌等),与宿主的生理特征(如皮脂分泌、角质代谢、免疫状态)及外部环境因素(如温度、湿度、pH 值)相互作用,共同构建的一个动态平衡系统[1]。
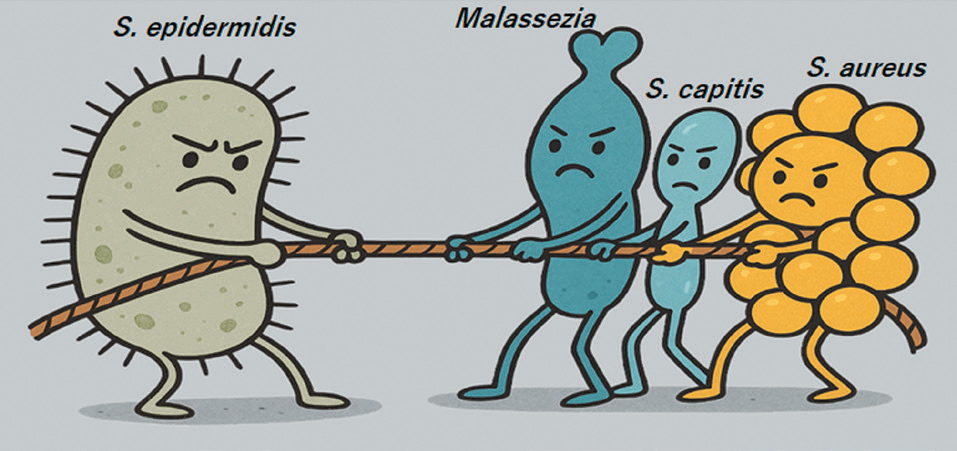
图 1. 头皮微生态平衡示意图
目前研究表明,头皮定植菌群主要包括葡萄球菌属(Staphylococcus spp.)、丙酸杆菌属(Propionibacterium spp.)和马拉色菌属(Malassezia spp.),微生物细胞密度约为 103–105 CFU/mm²[2]。这些微生物通过利用宿主代谢产物形成竞争与共生关系,并对环境做出适应性调节,从而维持头皮的稳态。
类似于肠道菌群,头皮菌群可依据对宿主的作用分为三类:(1)有益菌(beneficial bacteria):对头皮屏障、免疫及健康状态产生正面作用的菌种;(2)有害菌(pathogenic bacteria):与头屑、炎症甚至感染相关的菌种;(3)摇摆菌(opportunistic/commensal bacteria):根据环境与宿主状态,可能产生有益或有害作用的菌种。
头皮微生态平衡(scalp microecological balance)并不意味着“有益菌数量必须多于有害菌”,而是菌群多样性与稳定性保持在动态均衡状态。在这一平衡中,有益菌及摇摆菌对有害菌形成生态“牵制”,使其不至于异常增殖。当这种平衡被打破(dysbiosis)——例如特定致病菌(如马拉色菌)大量繁殖——便可能导致头屑、炎症、瘙痒等头皮问题(图1)。
头皮常见“有益菌”及“有害菌”
(1)表皮葡萄球菌(Staphylococcus epidermidis)
——典型”有益菌”
表皮葡萄球菌属于凝固酶阴性葡萄球菌(CoNS),是人类皮肤和黏膜的常驻菌群,通常不产生外毒素,具有良好的共生性[3]。其作用包括:
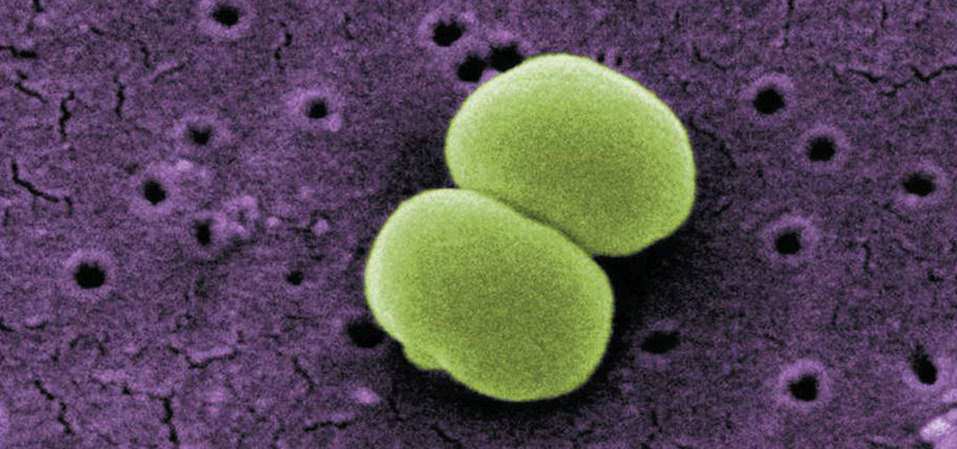
图2. 表皮葡萄球菌
(图片来源:Public Health Image Library (PHIL), ID#:10041)
• 代谢皮脂、维持酸性屏障:分解皮脂中的甘油三酯产生短链脂肪酸,维持皮肤弱酸性环境,抑制病原菌增殖[4-5]。
• 产生抗菌物质:分泌多种抗菌肽(epidermin、epilancin 等)及酚溶性调控蛋白(PSMs),抑制金黄色葡萄球菌、化脓性链球菌等致病菌定植[6-8]。
• 调节免疫与抗炎:通过脂磷壁酸(LTA)激活角质形成细胞表面的Toll样受体2(TLR2)产生miRNA-143,下调促炎反应,并促进 β-防御素生成[10-11]。
• 促进屏障功能:其分泌的鞘磷脂酶可促进角质层神经酰胺生成,增强皮肤保湿力[12]。
基于这些作用,表皮葡萄球菌在头皮上普遍被视为“有益菌”。
(2)金黄色葡萄球菌(Staphylococcus aureus)
——典型”有害菌”
与表皮葡萄球菌不同,金黄色葡萄球菌为凝固酶阳性葡萄球菌(CoPS),可利用纤维蛋白包裹自身,逃避免疫系统攻击[13-14]。其作用有:
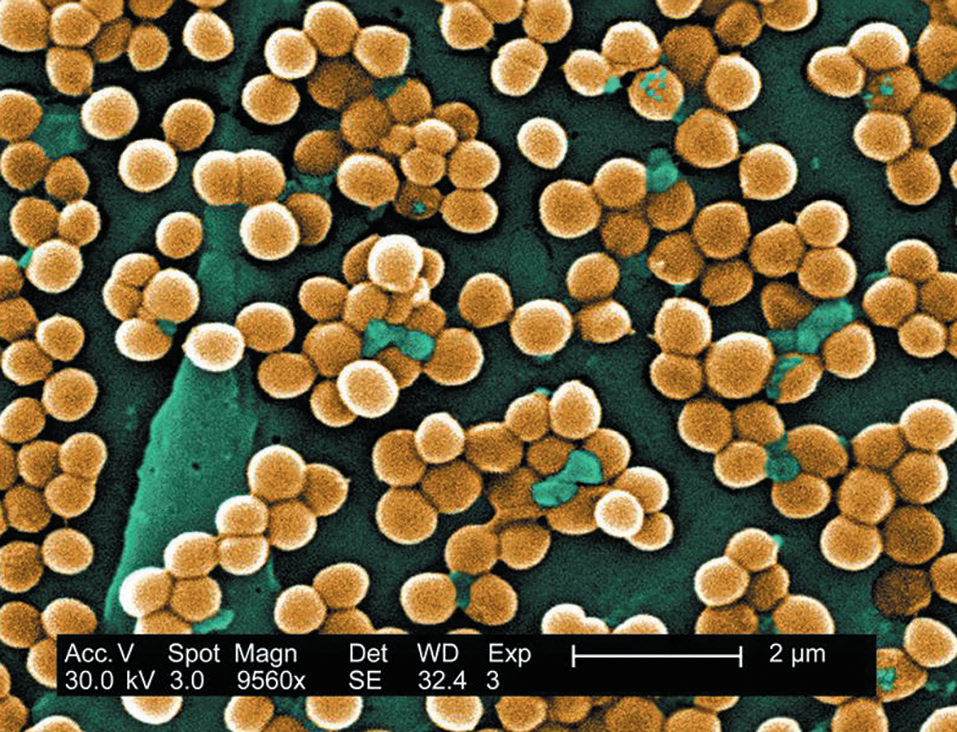
图3. 金黄色葡萄球菌
(图片来源:Public Health Image Library (PHIL), ID#:10046)
• 产毒致炎:可分泌 α-溶血素(α-toxin)破坏角质形成细胞,引发皮肤屏障受损和炎症[16];释放超级抗原(SEA/SEB、TSST-1)诱发严重局部炎症[17];通过Panton–Valentine leukocidin (PVL) 杀伤中性粒细胞,引发化脓性皮肤感染[18]。
• 免疫逃逸机制:通过D-丙氨酸修饰磷壁酸减弱自身净负电荷、分泌蛋白酶和溶菌素、产生纤溶酶葡萄球菌激酶 (SAK)等多重机制抵御宿主 CAMP(阳离子抗菌肽)杀伤,并削弱宿主防御反应[13]。
综上,金黄色葡萄球菌对宿主无明显益处,且有多重毒力因子,被普遍视为头皮“有害菌”。
(3)头葡萄球菌(Staphylococcus capitis)
——从“条件性致病菌”到潜在“有害菌”
头葡萄球菌与表皮葡萄球菌同属CoNS,理论上毒性较低,但其在头皮生态中的作用呈现争议性[10]:
头葡萄球菌
(图片来源:Wikipedia)
• 抗菌潜力有限:虽可分泌部分酚溶性调控蛋白(PSMs),对抗金黄色葡萄球菌和丙酸杆菌,但似乎抵抗能力不大[14]、且无明确抗菌肽产生证据[19-20]。
• 与头屑相关性强:多项研究发现,头屑人群中头葡萄球菌丰度显著升高,而表皮葡萄球菌丰度下降[21-23]。
• 潜在致病机制:头葡萄球菌诱导金黄色葡萄球菌毒力因子之一的 4,4′-二氢植物烯脱氢酶基因(4,4′-diapophytoene desaturase)高表达[22]。
因此,虽然头葡萄球菌在健康头皮中可能呈中性,但基于现有头屑相关证据,其在头皮护理语境中更多被归为“有害菌”。
(4)马拉色菌(Malassezia spp.)
——头皮屑“元凶”
马拉色菌属于好脂性真菌,是头皮最主要的真菌群之一,已确认有17 个菌种,其中约 10 种可在人类皮肤上分离[24]。已知其对头皮的影响如下:
马拉色菌
(图片来源:Public Health Image Library (PHIL), ID#:218)
• 头屑形成机制:头屑患者头皮上马拉色菌数量是正常人的 1.5–2.0 倍[25]。其通过带有LIP1基因的脂肪酶分解皮脂产生不饱和脂肪酸,刺激头皮引发角质细胞异常脱落,诱导IL-1β、TNF-α等炎症因子释放诱发炎症反应[26-28]。
• 多重皮肤疾病关联:可通过释放纳米级外泌体诱发炎症反应,加重特应性皮炎、银屑病,甚至导致毛囊炎[24]。
• 益处有限:虽可产生部分抗真菌的短链脂肪酸(如壬二酸),但其负面影响远大于潜在益处[24]。
因此,马拉色菌被广泛认定为头屑与脂溢性皮炎的关键“有害菌”。
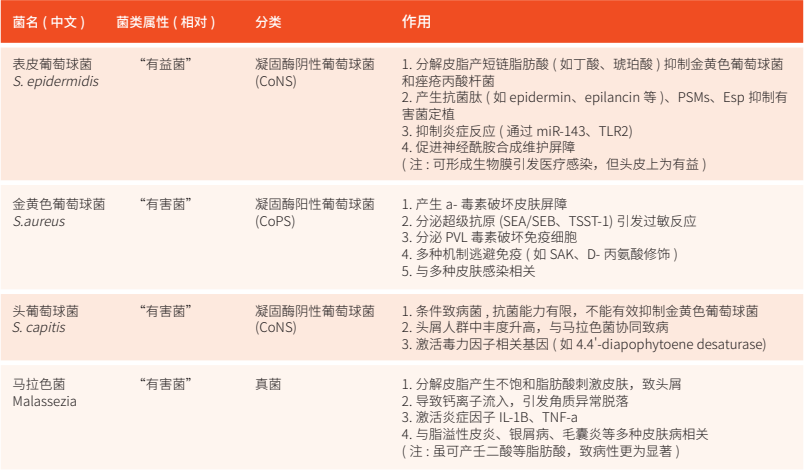
表 1. 头皮主要“有益菌”和“有害菌”
“抑恶扬善”式头皮护理策略展望
本文对多种头皮常见的“有益菌”与“有害菌”进行了梳理(见表1)。必须指出的是:头皮微生物群与宿主之间的关系极为复杂,不仅受身体部位、年龄及宿主易感性影响,甚至同一种菌在不同部位的作用也可能完全不同。例如,被称为“摇摆菌”的痤疮丙酸杆菌(Propionibacterium acnes)是健康头皮上丰度最高的菌群之一,但由于其在头皮上并不易引发痤疮,本文暂未做详细阐述。关于“有益菌”与“有害菌”的分类,学界并无绝对标准,这更凸显了精准干预与科学管理的重要性。
针对头皮问题,头皮护理理念应从单纯“抑菌”转向“抑恶扬善”:在抑制致病菌的同时扶持有益菌群,通过菌群间的正向竞争与调节,恢复并维持头皮健康的微生态平衡。然而,头皮护理技术的完善仍需临床研究支持。未来应更多考虑皮肤微生物菌群的个体解剖学差异,并结合宿主的易感性、性别、年龄、季节及种族等多维因素,建立大规模数据库,以明确菌群变化对皮肤稳态的直接影响并加深对因果关系的理解。这不仅有助于构建更精准的干预模型,也为个体化护理产品的研发提供了基础。
随着头皮健康理念的普及和微生态研究的深入,“抑恶扬善式菌群平衡策略”有望成为未来头皮护理领域的核心技术支柱。 该策略通过选择性抑制致病菌、促进益生菌的协同生长,可在油脂调控、头屑改善、炎症缓解、毛囊环境优化、防脱发及促进生发等多个维度提供长期支持。
更具前景的是,随着头皮微生态测序(NGS)技术、AI 数据建模与个性化配方平台的成熟,“抑恶扬善”策略有望与定制化护理产品深度融合。未来,品牌或医疗机构可依据个体的“菌群指纹”设计个性化干预方案,涵盖从基础清洁、精华护理到营养补充的全链路管理,推动头皮护理模式从“共性护理”走向“精准干预”。这种以科学验证与个性化解决方案为核心的头皮管理框架,或将推动行业从消费级洗护产品进一步迈向医研结合、预防干预的新阶段。
作者:王放放1, 林娴婷1, 杨建中1,2
1美研创新株式会社,2 株式会社 O and R
参考文献:
[1] O'Neill AM, Gallo RL. Host-microbiome interactions and recent progress into understanding the biology of acne vulgaris. Microbiome. 2018;6(1):177.
[2] 王丽等. 头皮微生物多样性与去头屑活性成分研究进展. 微生物学报. 2019; 46(10): 2772−2780.
[3] Cameron DR, Jiang JH, Hassan KA, et al. Insights on virulence from the complete genome of Staphylococcus capitis. Front Microbiol. 2015; 6:980.
[4] Wang Y, Kuo S, Shu M, Yu J, Huang S, Dai A, Two A, Gallo RL, & Huang CM. Staphylococcus epidermidis in the human skin microbiome mediates fermentation to inhibit the growth of Propionibacterium acnes: implications of probiotics in acne vulgaris. Applied microbiology and biotechnology. 2014; 98(1), 411–424.
[5] Keshari S, Balasubramaniam A, Myagmardoloonjin B, Herr DR, Negari IP, & Huang CM. Butyric Acid from Probiotic Staphylococcus epidermidis in the Skin Microbiome Down-Regulates the Ultraviolet-Induced Pro-Inflammatory IL-6 Cytokine via Short-Chain Fatty Acid Receptor. International journal of molecular sciences. 2019; 20(18), 4477.
[6] Torres Salazar, Benjamin O et al. “Commensal production of a broad-spectrum and short-lived antimicrobial peptide polyene eliminates nasal Staphylococcus aureus.” Nature microbiology. 2024; 9(1): 200-213.
[7] Cogen AL, Nizet V, & Gallo RL. Skin microbiota: a source of disease or defence?. The British journal of dermatology. 2008; 158(3), 442–455.
[8] Cogen, Anna L et al. Staphylococcus epidermidis antimicrobial delta-toxin (phenol-soluble modulin-gamma) cooperates with host antimicrobial peptides to kill group A Streptococcus. PloS one. 2010; 5,1 e8557. 5 Jan.
[9] Iwase T, Uehara Y, Shinji H, Tajima A, Seo H, Takada K, Agata T, Mizunoe Y. Staphylococcus epidermidis Esp inhibits Staphylococcus aureus biofilm formation and nasal colonization. Nature. 2010; 465:346–349.
[10] Xia, Xiaoli et al. Staphylococcal LTA-Induced miR-143 Inhibits Propionibacterium acnes-Mediated Inflammatory Response in Skin. The Journal of investigative dermatology. 2016; 136(3): 621-630.
[11] Lai, Yuping et al. Activation of TLR2 by a small molecule produced by Staphylococcus epidermidis increases antimicrobial defense against bacterial skin infections. The Journal of investigative dermatology. 2010; 130(9): 2211-21.
[12] Y. Zheng et al. Commensal Staphylococcus epidermidis contributes to skin barrier homeostasis by generating protective ceramides. Cell Host Microbe. 2022; 30(3):301-313.e9
[13] Ryu, S, Song, PI, Seo, CH, Cheong, H, & Park, Y. Colonization and infection of the skin by S. aureus: immune system evasion and the response to cationic antimicrobial peptides. International journal of molecular sciences. 2014;15(5), 8753–8772.
[14] Parlet, Corey P et al. Commensal Staphylococci Influence Staphylococcus aureus Skin Colonization and Disease. Trends in microbiology. 2019; 27(6):497-507.
[15] Chen YE et al. Skin microbiota-host interactions. Nature. 2018; 553 (7689):427–436.
[16] Berube BJ, Bubeck Wardenburg J. Staphylococcus aureus α-toxin: nearly a century of intrigue. Toxins (Basel). 2013;5(6):1140-1166.
[17] Laouini D, Kawamoto S, Yalcindag A, et al. Epicutaneous sensitization with superantigen induces allergic skin inflammation. J Allergy Clin Immunol. 2003;112(5):981-987.
[18] Shallcross LJ, Fragaszy E, Johnson AM, Hayward AC. The role of the Panton-Valentine leucocidin toxin in staphylococcal disease: a systematic review and meta-analysis. Lancet Infect Dis. 2013;13(1):43-54.
[19] Flowers L, Grice EA. The Skin Microbiota: Balancing Risk and Reward. Cell Host Microbe. 2020;28(2):190-200.
[20] O'Neill AM, Nakatsuji T, Hayachi A, et al. Identification of a Human Skin Commensal Bacterium that Selectively Kills Cutibacterium acnes. J Invest Dermatol. 2020;140(8):1619-1628.e2.
[21] Grimshaw SG, Smith AM, Arnold DS, Xu E, Hoptroff M, Murphy B. The diversity and abundance of fungi and bacteria on the healthy and dandruff affected human scalp. PLoS One. 2019;14(12):e0225796.
[22] Hu P, Henry J, Tiesman JP, et al. Scalp microbiome composition changes and pathway evaluations due to effective treatment with Piroctone Olamine shampoo. Int J Cosmet Sci. 2024;46(3):333-347.
[23] Xu Z, Yang F, Xiang B, et al. Abnormal Microbial Amino Acid Metabolism and Activated Pathogenesis in Scalp with Dandruff. J Invest Dermatol. 2025;145(7):1823-1826.e7.
[24] Vijaya Chandra SH, Srinivas R, Dawson TL Jr, Common JE. Cutaneous Malassezia: Commensal, Pathogen, or Protector?. Front Cell Infect Microbiol. 2021;10:614446.
[25] McGinley KJ, Leyden JJ, Marples RR, et al. Quantitative microbiology of the scalp in non-dandruff, dandruff, and seborrheic dermatitis. Journal of Investigative Dermatology.
1975;64(6): 401-405
[26] DeAngelis YM, Saunders CW, Johnstone KR, et al. Isolation and expression of a Malassezia globosa lipase gene, LIP1. J Invest Dermatol. 2007;127(9):2138-2146.
[27] Y Katsuta, T Iida, S Inomata, and M Denda. Unsaturated Fatty Acids Induce Calcium Influx into Keratinocytes and Cause Abnormal Differentiation of Epidermis. J Invest Dermatol. 2005;124(5):1008–1013.
[28] Sparber F, De Gregorio C, Steckholzer S, et al. The Skin Commensal Yeast Malassezia Triggers a Type 17 Response that Coordinates Anti-fungal Immunity and Exacerbates Skin Inflammation. Cell Host Microbe. 2019;25(3):389-403.e6.
来源:荣格-《国际个人护理品生产商情》
原创声明:
本站所有原创内容未经允许,禁止任何网站、微信公众号等平台等机构转载、摘抄,否则荣格工业传媒保留追责权利。任何此前未经允许,已经转载本站原创文章的平台,请立即删除相关文章。


