 供需大厅
供需大厅
 登录/注册
登录/注册 供应商登录
供应商登录










荣格工业资源APP
了解工业圈,从荣格工业资源APP开始。
欢迎来到荣格工业资源网!

供需大厅

登录/注册
公众号

更多资讯,关注微信公众号
小秘书

更多资讯,关注荣格小秘书
邮箱
您可以联系我们 info@ringiertrade.com

电话
您可以拨打热线
+86-21 6289-5533 x 269
建议或意见
+86-20 2885 5256
顶部

荣格工业资源APP
了解工业圈,从荣格工业资源APP开始。



如果把神经退行性疾病比作一座日渐崩塌的能量宫殿,线粒体 DNA(mtDNA)无疑是那块最易破裂、却至关重要的地基。近年来,一种不为大众熟知却被科学界寄予厚望的策略正在悄然铺开——修复线粒体内部的基因结构,重新点燃细胞的能量引擎,挽救那些缓慢死亡的神经元。
从帕金森病(PD)、阿尔茨海默病(AD),到亨廷顿舞蹈病(HD)和肌萎缩侧索硬化症(ALS),研究者们正试图绕过传统蛋白靶点干预路径,深入线粒体基因组内部,通过精准的碱基编辑技术,抹去那些导致功能失调的突变「笔迹」,从根源干预疾病发生机制。
今天,《国际医药商情》这篇文章将梳理这一领域的最新研究动态与技术进展,尤其聚焦 DdCBE、TALED、mitoZFDs 等新一代编辑工具的应用潜力,并评估其在治疗神经退行性疾病中的可行性、瓶颈与未来走向。
Part 1
打破线粒体封闭体系,基因编辑的新突围
在基因编辑技术持续高歌猛进的当下,线粒体基因却一直是被「遗忘的角落」。
原因不难理解——线粒体具有独立于细胞核的基因组,不仅结构紧凑、遗传规则特殊(主要为母系遗传),更关键的是,它被两层脂质双层膜紧密包裹,仿佛一座分子层级上的孤岛,拒绝大部分外源编辑工具的进入。
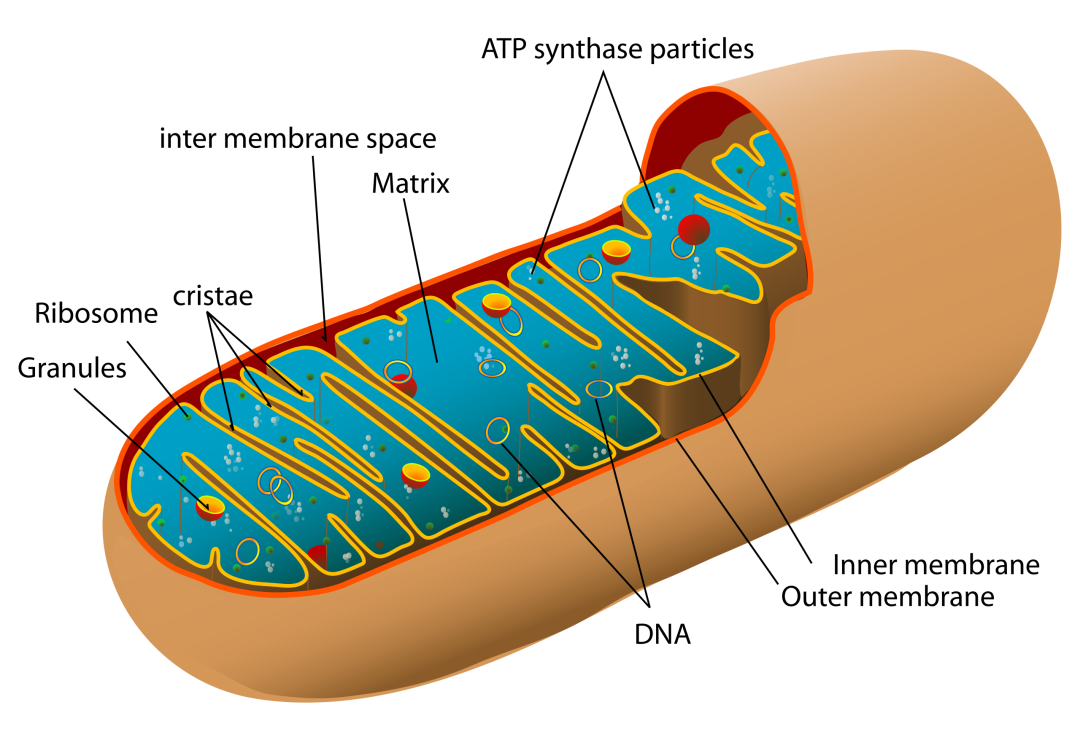
最早一批尝试突破这一「封闭体系」的,是人工核酸酶类工具,包括线粒体锌指核酸酶(mitoZFN)与线粒体转录激活因子样效应物核酸酶(mitoTALEN)。这两种技术都基于「蛋白识别 + 核酸酶切割」的基本逻辑,通过添加线粒体靶向序列(MTS),使得这些蛋白编辑工具可以进入线粒体内进行特定位点的双链 DNA 切割。
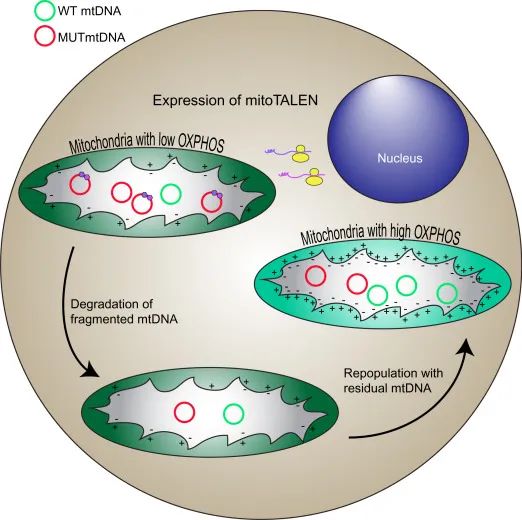
然而,这种「定点清除突变 mtDNA」的策略,在实际应用中面临两个困局:其一是编辑效率低,难以建立持续的治疗效果;其二是仅能切除突变,无法进行精确的碱基替换修复,对一些致病突变而言,仍显力不从心。
真正改变局势的,是基于脱氨酶的线粒体碱基编辑技术(mitochondrial base editors)的出现。
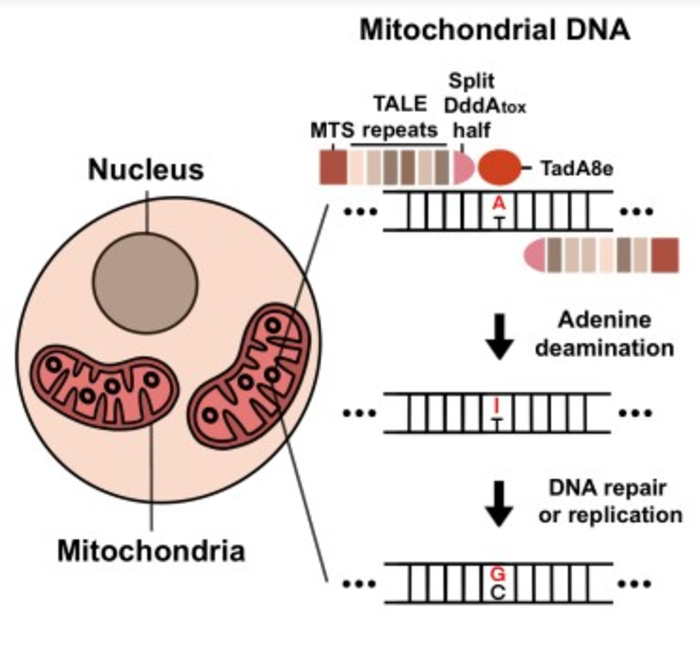
其中最具代表性的,是 2020 年由美国博德研究所(Broad Institute)的 David R. Liu 团队与 Joseph D. Mougous 团队开发的DdCBE(double-stranded DNA cytosine base editor,双链 DNA 胞嘧啶碱基编辑)。
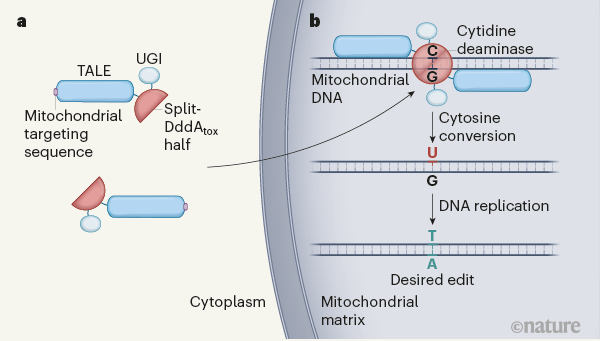
该系统将一对 DddA 脱氨酶亚基分别连接至 TALE DNA 识别模块上,并配合尿嘧啶 DNA 糖基化酶抑制剂(UGI)构建而成,能够在不切断 DNA 双链的情况下,实现精准的 C-to-T 点突变编辑。这一系统不依赖向线粒体引入 gRNA,因此规避了 CRISPR-Cas9 无法进入线粒体的天然限制。
继 DdCBE 之后,研究者们进一步构建出 mitoZFDs(线粒体锌指脱氨酶)与 TALEDs(转录激活因子样效应物腺嘌呤脱氨酶),分别实现了更小体积、更高包装效率的碱基编辑,以及首个 A-to-G 碱基替换操作。这意味着,研究者已经能够在 mtDNA 中覆盖更广泛的点突变类型,并有机会对绝大多数线粒体遗传病位点实现理论上的编辑可及性。
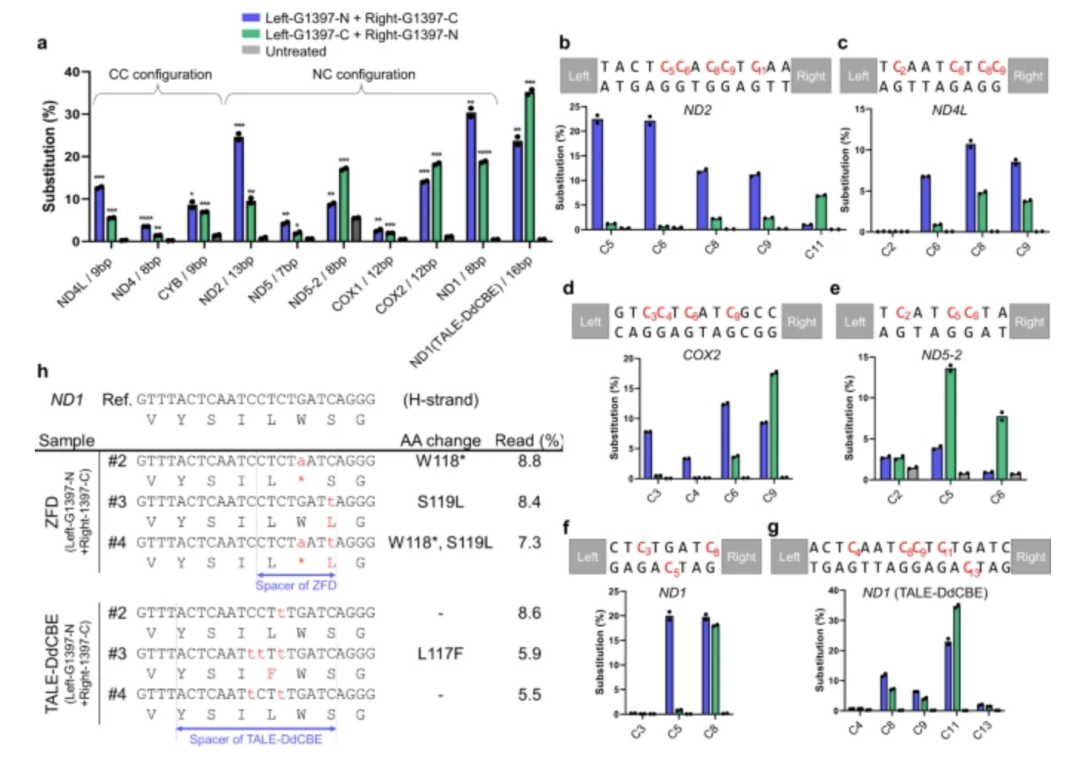
更进一步,mitoBEs(线粒体双脱氨酶系统)将 nickase 酶与脱氨酶组合,甚至可以同时实现 A-to-G 与 C-to-T 的双向编辑,在模型细胞中展现出更高效率、更低脱靶的特性。相比于传统的基因敲除或突变拮抗技术,这些不切断双链的「软性编辑」手段无疑更具安全性,尤其适合应用于神经系统这种对氧化应激与突变高度敏感的组织。
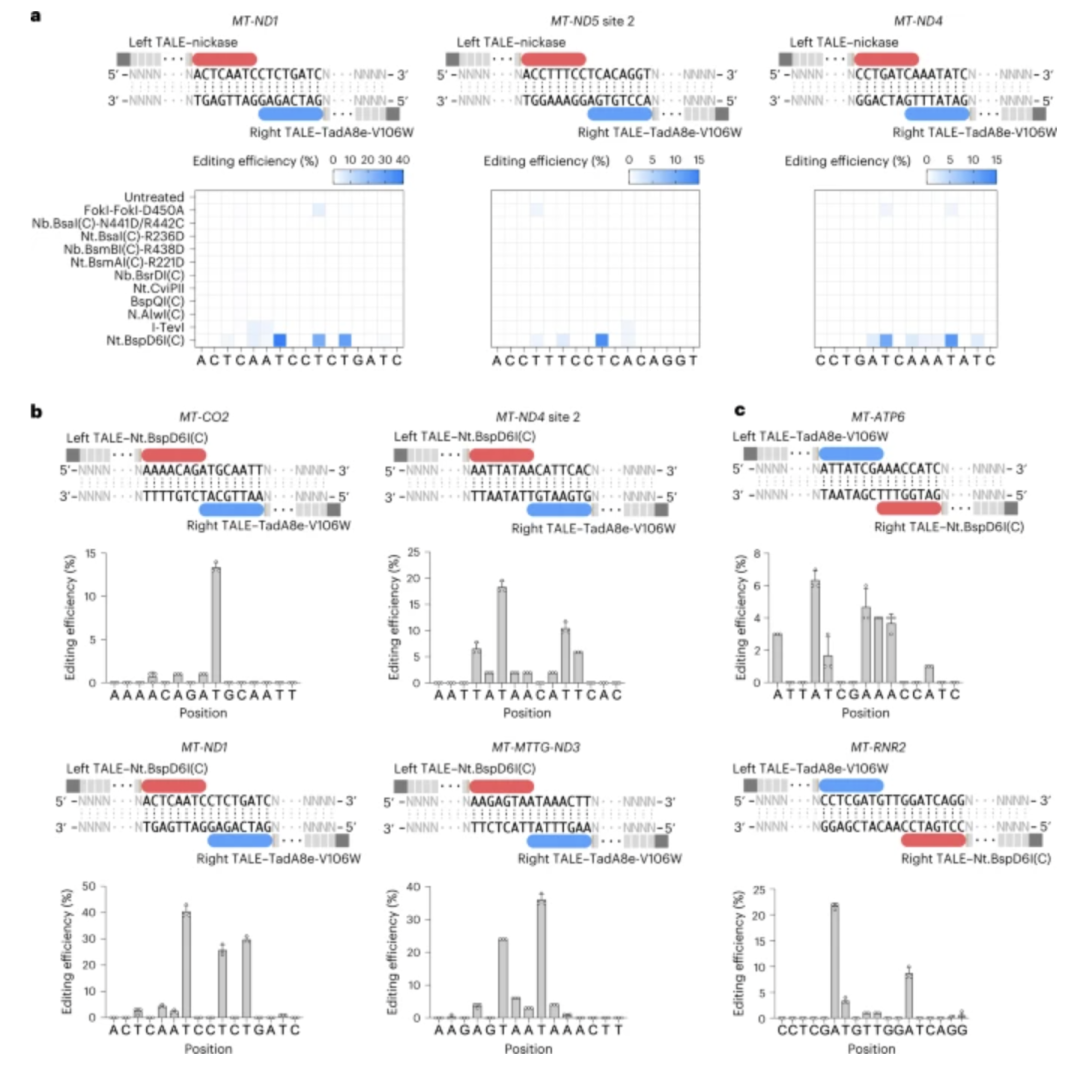
技术上的突破使得线粒体基因编辑从不可能走向可能,但另一个问题也随之浮现——如何将这些复杂的蛋白编辑系统「精准投递」进细胞线粒体内部?
目前,研究者们主要依赖三种策略——其一是 MTS 肽段引导,通过自然的线粒体蛋白转运通路将编辑酶引入胞器内;其二是病毒载体系统(如 AAV),借助病毒自身的跨膜运输能力,将编辑组件送达特定组织线粒体中;其三是新兴的非病毒递送工具,如脂质纳米粒子(LNP)与类病毒颗粒(VLP),正在为更大体积的编辑系统探索更安全的交付路径。
不过,无论是哪种方式,目前最核心的瓶颈仍在「有效负载与线粒体靶向效率」之间的平衡问题。
AAV 的包装容量有限,不足以一次性负载完整的 DdCBE 或 TALED;而 LNP 系统虽然容量充裕,但线粒体内吞能力极弱,仍在实验室早期探索阶段。即便如此,研究者仍然通过如「环状 RNA 编码的 mitoBEs」等方式,试图在编辑效率、系统稳定性与靶向特异性之间找到最佳解法。
Part 2
线粒体 DNA 变异
驱动神经退行性疾病的 4 种典型路径
在神经退行性疾病的发病链条中,线粒体 DNA 的角色一直被低估。
如今,随着超高通量测序与单细胞技术的发展,研究者开始逐步揭开 mtDNA 突变在不同类型神经元死亡过程中的真实面貌。
帕金森病(PD)、阿尔茨海默病(AD)、亨廷顿病(HD)、肌萎缩侧索硬化症(ALS)——这四种最常见、最具社会负担的 NDD,其背后都藏有线粒体功能障碍的痕迹,而线粒体 DNA 突变,正是那个引爆多米诺骨牌的起始点。
帕金森病(PD):
线粒体拷贝数调控失衡与突变积聚
帕金森病的核心病理是黑质多巴胺能神经元的进行性退化死亡。尽管 α-突触核蛋白(α-syn)聚集、Lewy 小体形成是诊断依据,但其根本驱动力之一却是线粒体功能崩溃。
早在 2010 年前后,就有研究指出,帕金森患者黑质区域中 mtDNA 的大量缺失与拷贝数下降密切相关。新近的研究进一步明确——不是点突变本身,而是 mtDNA 拷贝数调节机制的失灵,才是驱动线粒体功能恶化的关键因子。挪威卑尔根豪克兰大学医院神经系统医学中心的 Christian Dölle 等人的研究显示,mtDNA 拷贝数在 PD 患者神经元中显著下降,导致野生型 mtDNA 不足以维持线粒体正常呼吸链功能,最终导致能量危机和细胞凋亡。
此外,多个核编码线粒体维护基因(如 POLG、c10orf2 和 MPV17)也在 PD 患者中发现功能障碍,暗示 PD 可能是一种「核-线粒体互作紊乱型」疾病。在甲基化水平方面,PD 患者 D-loop 区域表现出明显去甲基化,可能干扰 mtDNA 复制起始,进一步加剧突变与缺失。
阿尔茨海默病(AD):
突变频率升高与表观遗传调控异常
阿尔茨海默病的线粒体路径略有不同。研究者发现,AD 患者海马体区域的 mtDNA 突变频率显著升高,尤其是 COX III 基因中的 T→C 突变,与线粒体氧化磷酸化能力下降高度相关。英国纽卡斯尔大学的 Kim J. Krishnan 等人的研究指出,COX 缺陷神经元在 AD 患者中显著增加,而这一缺陷可追溯至 mtDNA 编码亚基的突变。
除了突变,线粒体 DNA 的甲基化模式也发生显著变化。西班牙贝尔维奇大学医院的 Marta Blanch 团队在 AD 早期患者的嗅皮质与 D-loop 区域中观察到明显的 CpG 与非 CpG 位点甲基化升高;另有研究指出,在 APP/PS1 小鼠模型中,D-loop 区域甲基化下降与 mtDNA 拷贝数减少并存,暗示其在疾病早期即已介入。
这些变化并非孤立事件,而可能通过 mt-ncRNA(线粒体编码或核编码非编码 RNA)参与核-线粒体之间的双向信号调控,进一步干扰细胞能量代谢与凋亡路径。
亨廷顿病(HD):
突变负荷与基因表达协同破坏
HD 是一种由 HTT 基因 CAG 重复扩增所致的单基因常染色体显性遗传病,但 mtDNA 突变在其发病过程中扮演着协同作用。
研究显示,HD 患者全基因组 mtDNA 中 13 个蛋白编码区均可检测到不同程度的突变异质性,且其水平与患者运动与认知能力恶化成负相关。康奈尔大学的 Yiqin Wang 等人在长期随访样本中发现,mtDNA 突变负荷越高,HD 患者表现出更明显的认知退化。
此外,R6/2 与 STHdhQ111 这两种 HD 小鼠模型也分别表现出 mtDNA 拷贝数下降与损伤积累,进一步佐证了线粒体通路在 HD 发病中的核心地位。研究发现,tRNALeu 基因中的 m.3244 突变在 HD 样本中高度流行。鉴于 mitoTALEN 已在其他 tRNA 突变模型(如tRNAAla m.5024C>T突变小鼠)中证明能够显著降低突变 mtDNA 水平,未来针对 tRNALeu 位点开发的线粒体碱基编辑器可能成为缓解或延缓 HD 进展的潜在治疗策略。
肌萎缩侧索硬化症(ALS):
线粒体甲基化改变与补偿机制推演
ALS 的病因更加复杂,涵盖 SOD1、C9orf72、FUS 等多种核基因突变。然而,mtDNA D-loop 区甲基化水平与拷贝数呈显著负相关的发现,为理解 ALS 中的线粒体代谢失衡提供了新线索。
具体而言,在 SOD1 与 C9orf72 突变携带者中,尽管 mtDNA 拷贝数增加,但其调控区 D-loop 甲基化水平却下降,提示细胞可能存在一种「线粒体能量补偿机制」,试图通过增加 mtDNA 数量来对抗线粒体损伤。这种调节过程一旦被打破,便可能转为病理性代偿,加速运动神经元退化。
目前,ALS 的基因治疗已进入临床阶段,但大多聚焦于核基因突变干预。若能结合线粒体基因编辑工具,在 mtDNA 层面进行精细化调控,或将为 ALS 治疗打开新窗口。
Part 3
线粒体编辑技术的现实挑战与未来可能
在科研论文的页脚,线粒体基因编辑的每一项突破都写得令人振奋,但在转化医学的时间轴上,这些技术距离真正「落地」仍有一段崎岖的距离。
尤其是在神经退行性疾病这样复杂且高度异质的疾病谱中,如何从动物模型的「可编辑」跨越到人类临床的「可治疗」,是摆在所有研究者面前最沉重的命题。
第一个难题,是递送系统的「工程化瓶颈」。
目前被广泛使用的 AAV 载体虽然已经在某些罕见病基因疗法中获得 FDA 批准,但其最大包装能力小于5 kb,这对于体积庞大的 DdCBE 或 TALED 而言已接近极限。
更复杂的是,线粒体靶向需要在原有基础上增加MTS等定向标签,使得「装载困难」的问题愈发严峻。即便通过双载体系统、Split-Intein 拼接等手段勉强解决结构问题,递送效率、稳定表达与组织特异性表达仍难以兼顾。研究者正尝试以类病毒颗粒(VLP)、LNP 甚至自组装蛋白纳米笼为基础,构建全新的线粒体递送平台,但距离临床应用仍属前沿探索阶段。
第二个瓶颈,是脱靶效应的控制。
与核基因编辑不同,线粒体缺乏 NHEJ 或 HDR 等高效的双链断裂修复机制,主要依赖碱基切除修复(BER)等相对有限的修复途径,一旦发生非特异性剪切,往往难以有效修复,几乎意味着不可逆的功能丧失。此前关于 DdCBE 的研究曾指出,即使在未进行 DNA 靶向的情况下,其 DddAtox 半酶分子也可能在细胞中自发重组,导致随机 C→T 突变,甚至波及核 DNA。为此,研究者开发了 NES-DdCBE(携带核输出信号)等策略,试图降低核定位错误,并通过工程化改造 DddAtox 结构、引入天然抑制元件等方式抑制「野性表达」的风险。
但更棘手的问题在于,长期表达带来的线粒体突变积累效应,可能在未来数月甚至数年后才显现为系统性病理。这意味着临床试验必须进行超长周期的追踪与安全性验证,对研究资源和伦理体系提出极高要求。
第三个现实障碍,是对 mtDNA 动态变化的系统认知尚不足。
与固定突变位点不同,线粒体内的 mtDNA 呈现出高度的异质性与动态重组趋势。在某些组织中,突变 mtDNA 即使仅占 20%也能触发病变,而在另一些组织中,可能需要超过 60%才会显现功能障碍。这种所谓「阈值效应」意味着,编辑策略不应仅追求全清或全修,而应建立在组织特异、剂量可控、干预时间窗明确的基础上。因此,未来的发展方向之一,是通过数学建模与系统生物学工具,模拟个体异质性下的最佳编辑剂量、干预时机与突变替代比例,实现更加精细化的治疗方案制定。
第四个现实挑战,是伦理监管的约束与未来风险。
UNESCO 和 WHO 等国际组织对基因编辑监管日趋严格。2021 年,WHO 发布首个全球基因组编辑治理框架,强调技术应用必须「安全、有效且合乎伦理」。线粒体基因编辑面临种系遗传风险——由于线粒体 DNA 通过母系遗传,编辑后的遗传修饰可能传递给后代。目前仅英国合法化某些线粒体替代技术,且严格限制在预防严重遗传疾病范围内。安全性方面,最新研究证实即便先进的 DdCBE 系统也存在核基因组脱靶编辑风险,长期影响仍需深入评估。WHO 国际共识是,在安全性和有效性充分验证前,不应进行可遗传的人类基因组编辑。
Part 4
结语
下一个突破窗口,会在中国吗?
在全球线粒体编辑研究竞速中,中国正在迅速崛起。中国在线粒体基因编辑技术方面已有重要进展,相关研究得到国家重点研发计划等重要资助支持。不过,在创新性突破方面仍需提升。
在神经退行性疾病负担方面,中国正面临巨大挑战。
首都医科大学附属北京天坛医院冯涛团队基于 2021 年全球疾病负担研究(GBD 2021)的数据,预测了 2050 年全球 195 个国家和地区的帕金森病患病率,公布在《英国医学杂志》。其中预计到 2050 年,中国将成为帕金森病患病率最高的国家(583 例/10 万人)。
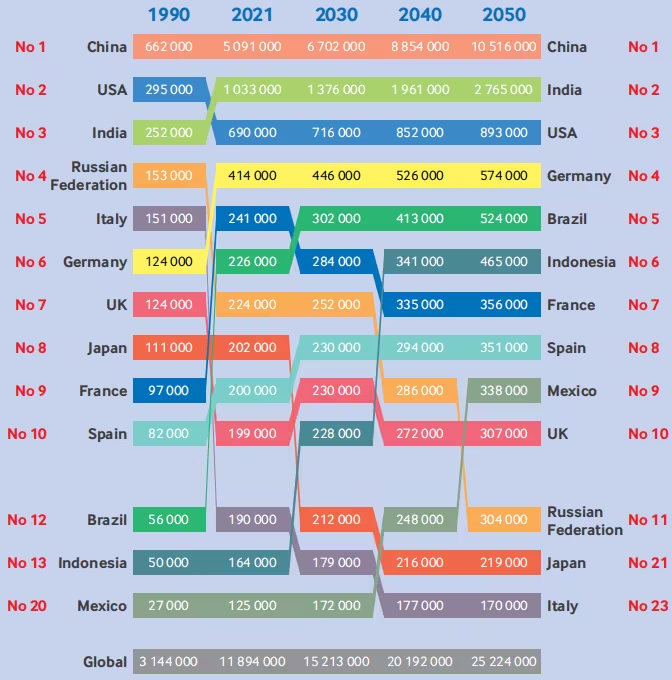
根据《General Psychiatry》期刊 2022 年发布的《中国阿尔茨海默病报告》,截至 2019 年,中国 60 岁及以上人群中,阿尔茨海默病患者约为 983 万人 。
中国学者在线粒体碱基编辑技术方面已有重要贡献,特别在神经退行性疾病研究领域,中国论文产出数量已位居世界第二位,近十年年复合增长率高达 20.01%。
以上海为代表的地区已出台专门的基因治疗产业发展行动方案(2023-2025 年),推动基因治疗科技创新与产业发展。

线粒体基因编辑确实是一项长期主义工程,其面对的是高度异质、进展缓慢但不可逆的神经系统疾病,对安全性、持久性、可控性要求极高。然而,正是这种对技术精度的极致要求,可能为中国在这一前沿领域实现突破提供机会。
在全球竞争中,时间窗口有限但机遇真实。若能在技术创新、监管完善、产业转化等方面协同发力,线粒体基因编辑技术有望成为中国生物医药产业在国际舞台上的重要突破口,为全球数以千万计的神经退行性疾病患者带来新的治疗希望。
参考资料:
Ye, H., Song, Y., Wang, W., Shi, J., & Chen, X. (2025). Mitochondrial DNA editing: Key to the treatment of neurodegenerative diseases. Genes & Diseases, Volume 12, Article 101437.
Blanch, M., Mosquera, J. L., Ansoleaga, B., et al. (2016). Altered mitochondrial DNA methylation pattern in Alzheimer disease-related pathology and in Parkinson disease. Acta Neuropathologica Communications, 4(1), 24.
Doxzen, K. W., & Huang, M. (2022). Extensive off-target editing by cytosine base editors in nuclear DNA of human cells. Nature, 610(7933), 416–424.
Zhou, Y., et al. (2022). China Alzheimer Report 2022. General Psychiatry, 35(1):e100751.
Yang, W., et al. (2024). Forecasting Parkinson’s disease burden to 2050. The BMJ, 388, e080952.
来源:国际医药商情
作者:John Xie


